村瀬 裕一 (佐藤研究室 博士課程)
Communications Biology誌
プレスリリース
Press release (English version)
医薬通信社記事
日本経済新聞記事
発表のポイント
早稲田大学 理工学術院(先進理工学研究科 生命医科学専攻 細胞骨格ロジスティクス研究室)の佐藤政充 教授らの研究グループは、岡山理科大学・東京大学との共同研究の成果として、染色体分配などの場面において、微小管を短縮化することで染色体を運搬する因子を発見しました。これまで分裂酵母細胞においては微小管を短縮化する因子としては別の因子が想定されていましたが、本研究では、想定外の因子が微小管の短縮化を実行することを立証しました。さらに、この因子を利用して人工的に微小管を短縮化することで染色体を運搬可能な分子機械を作製しました。これらの成果によって、これまでの染色体分配のイメージ図が大きく塗りかわる可能性があります。本研究成果は、Nature Portfolio journalsが提供するオープンアクセス・ジャーナル『Communications Biology』にて、2022年11月26日にオンラインで掲載されました。
(1)これまでの研究で分かっていたこと(科学史的・歴史的な背景など)
染色体分配は細胞分裂において最も重要な現象のひとつです。まず、微小管と呼ばれる繊維状の細胞骨格が紡錘体を形成し、これが染色体をつかまえます(図1)。その後、微小管は染色体を均等に2組に分配して、新しく作られる2個の細胞に運搬することで染色体分配が完了します。染色体分配が不均等になると、細胞死やがん化、あるいはダウン症候群などの先天性染色体異常の原因となると言われており、微小管が染色体を均等に2組に分配する分子メカニズムを解明することで、これらの疾患や症状の治療や予防のための基盤を作りたいと考えています。
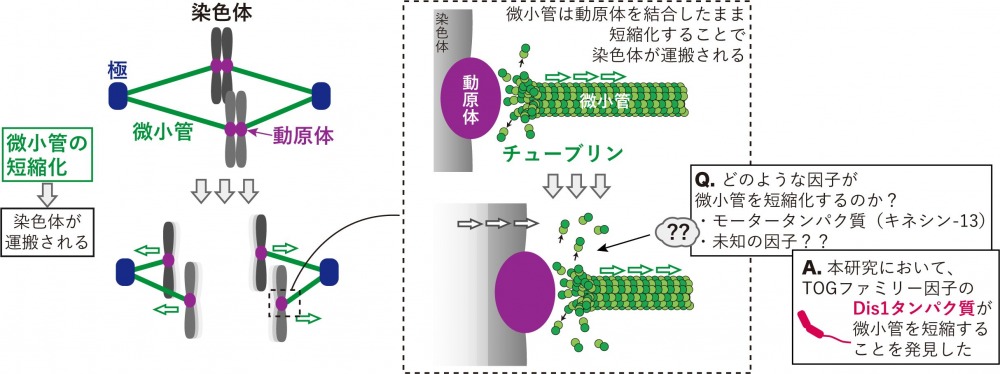
図1 細胞分裂における染色体分配
特に染色体がどのように運搬されるのかに注目してみると、まず、微小管が染色体の動原体と呼ばれる部位を結合します。その後、その結合部分において微小管が末端から短くなることで、染色体が運搬されていくと解釈されています(図1)。これまでの研究によると、微小管の短縮化を担う因子はキネシン-13と呼ばれるモータータンパク質の一種だとされています。
ところが、真核生物である分裂酵母(S. pombe)においては、キネシン-13をコードする遺伝子がゲノムに存在しません。そこで、少なくともこの生物ではキネシン-13以外の因子がその役割を担うと考えられてきました。具体的には、キネシン-13に類似した別のモータータンパク質であるキネシン-8が名指しされていました。しかしながら、分裂酵母でキネシン-8をノックアウト(遺伝子を人為的に除去)しても、細胞はまだじゅうぶん染色体を運搬することができます。そこで、分裂酵母ではキネシン-13でもキネシン-8でもない、まったく異なる未知の因子が微小管を短縮化して染色体を運搬すると私たちの研究室では考えていました。それでは、その未知の因子はいったい何でしょうか。これまでの私たちの実験結果から、ある意外な因子が微小管を短縮化するという予備的なデータが得られており、本研究ではそれを多角的に実証することにしました。
(2)今回の研究で新たに実現しようとしたこと、明らかになったこと
私たちが注目したのは、Dis1タンパク質です。Dis1は酵母からヒトまでの真核生物に広く保存された「TOGファミリー」に属するタンパク質(TOGタンパク質)です。分裂酵母には2種類のTOGタンパク質が存在し、ひとつはAlp14、もう一つがDis1と呼ばれます(図2)。TOGタンパク質は、一般的には微小管を伸長する因子(微小管ポリメラーゼ)として広く立証されています。したがって、今回の「Dis1が微小管を短縮する」という私たちの主張は、一般的な知見とは相容れない、正反対のものです。
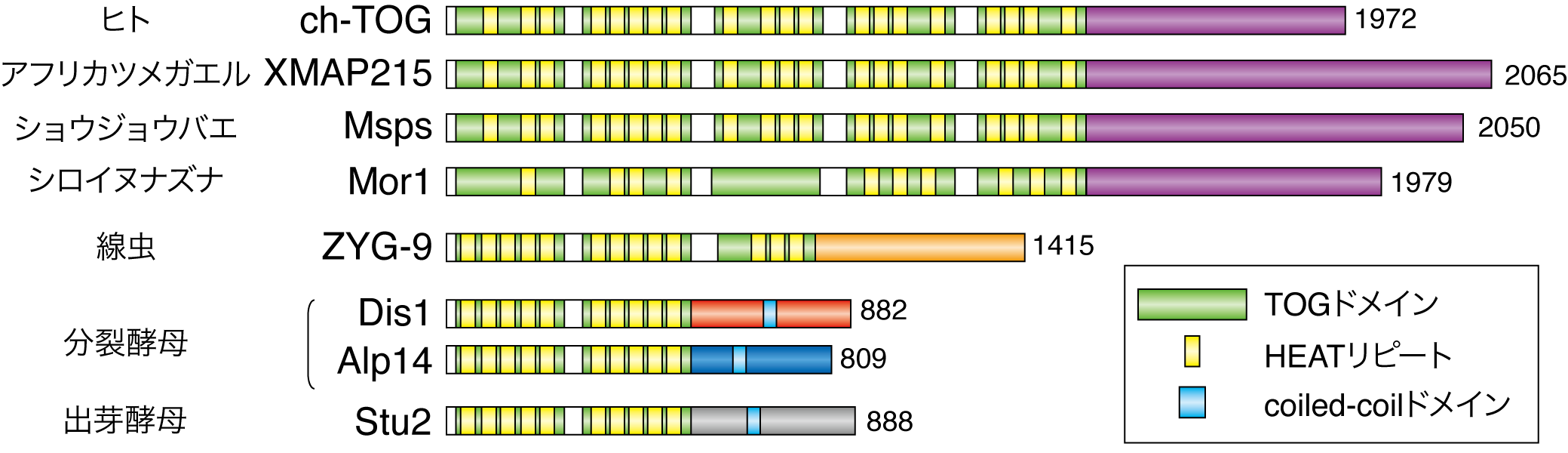
図2 各生物におけるTOGファミリーのタンパク質。分裂酵母のみ2種類のTOGタンパク質をもつことが特徴的である
そもそも、私たちは過去の研究成果において、減数分裂をおこなう細胞では微小管が特殊な機能を発揮して染色体を捕まえて引っ張ることを発見していました(Kakui et al. Nature Cell Biology, 2013、早稲田大学研究ニュース)。
当時すでに、TOGタンパク質であるDis1は微小管の伸長因子であると一般的に思われていましたが、私たちがdis1遺伝子をノックアウトした細胞を作製し観察したところ、予想に反して微小管が短縮化せず、染色体を運搬できないことが分かりました。そこから私たちは、Dis1タンパク質は微小管を短縮化する因子だとの仮説を立てました。とはいえ、このような定説に反することを主張するためには、dis1遺伝子のノックアウト細胞で起きていることを観察するだけでは不十分であり、Dis1タンパク質そのものに微小管を短縮化する活性があることを立証する必要があります。
そこで、岡山理科大学の濱田隆宏先生・東京大学の矢島潤一郎先生との共同研究として、試験管内での実証実験をおこないました。細胞から精製した微小管に対して、別途精製したDis1タンパク質を高濃度で添加したときに、微小管の動的な性質に変化が見られ、微小管の短縮が高頻度に起きるようになりました。このような試験管内での微小管動態アッセイから、Dis1には微小管を短縮化する活性があることの裏付けが取れたといえます。
ここで私たちは、この試験管内実験で得られた証拠をさらに補強するために、分裂酵母の細胞内に人工的な染色体運搬装置を構築することに挑戦しました。「Dis1は微小管を伸長するのではなく、逆に短縮する因子だ」という私たちの仮説が正しいならば、図3に示したように染色体のどこか1カ所(動原体ではない場所)にDis1だけを人工的に集積させて仮想的な動原体「バーチャル動原体」を作れば、Dis1の働きによって微小管が短縮化され、染色体運搬が起きるだろうと想定しました。
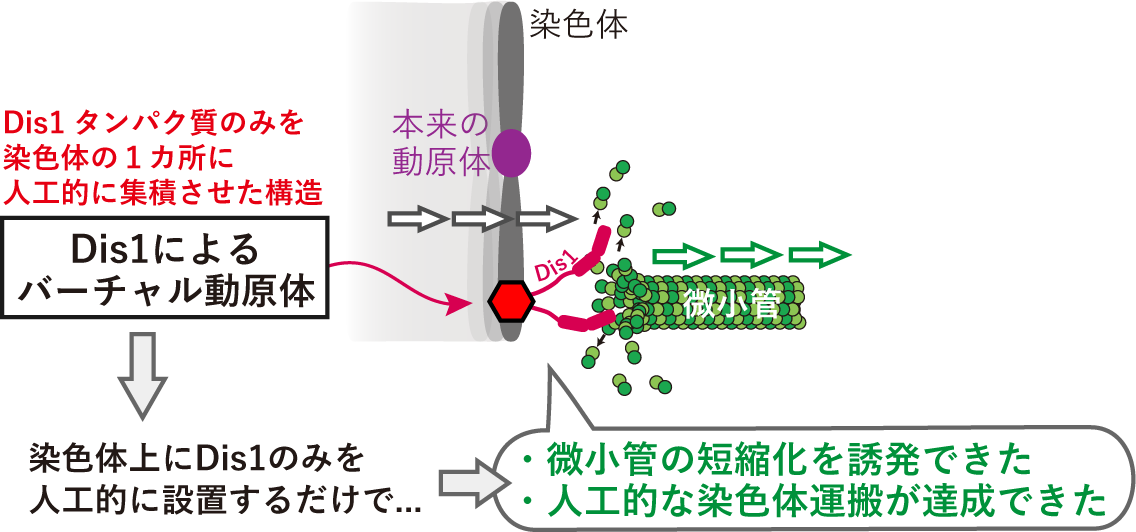
図3 Dis1を用いた人工的な染色体運搬装置の試作
この「バーチャル動原体」がうまく微小管をとらえて短縮化するかを試したところ、予想通り、染色体上に集積させたDis1が微小管と出会ったときに微小管を結合し、その微小管を短縮化しました。さらにその結果、染色体が運搬されることが実証できました(図3)。この「バーチャル動原体」にはDis1以外の動原体タンパク質は集まってこないことを確認しているので、ここで微小管をとらえてかつ短縮化する働きをもつのはすべてDis1によるものだといえます。しかも、Dis1のかわりに、もう一つのTOGタンパク質であるAlp14を人工的に集積させた場合には微小管の短縮化は起きませんでした。これらの結果は、微小管が染色体を結合して運搬するためには動原体のような多種多様なタンパク質は必要ではなく、Dis1さえあれば、ある程度達成できることを示しています。また、同じTOGタンパク質であってもAlp14(微小管を伸長する)とDis1(微小管を短縮化する)には正反対の機能があることも実証できました。真核生物の中でも分裂酵母だけがTOGタンパク質を2種類持っていましたが(図2)、両者にはこのような明確な使い分けがあったのです。
(3)研究の波及効果や社会的影響
教科書的には、微小管を短縮化するための因子はキネシン-13だとこれまで説明されてきましたが、本研究が示すように、定説をはるかに覆すようなTOGタンパク質Dis1もその役割を担うことが分かり、これまでの理解とは異なる細胞分裂の図式が見えてきました。さらに、Dis1さえ人工的に染色体上に集積させてしまえば、多数の構成因子によって成り立つ動原体の機能をある程度代替することができる、つまり「染色体と微小管を結合させて染色体を運搬する」ことが可能であることを実験で立証したことは、この分野の研究者にとってもサプライズとして受け止められています。
(4)今後の課題
一般的にはTOGタンパク質は微小管を伸長することが立証されているため、他のTOGファミリーとは異なりDis1のみが特殊な機能を獲得したのか、あるいは他のTOGファミリー因子も微小管の短縮化をおこなうのか、議論が続くものと思われます。また、本研究で試作した「Dis1による人工的な染色体運搬装置」をさらに洗練されたものにしていくことで、本来の動原体と同等の機能を担う「Dis1による人工動原体」が作製できると考え、これを生物学実験への適用、人工細胞の構築や医学的な応用に繋げていきたいと考えています。







